Промежуточный мозг. Таламус
Как и любой другой орган мозга, таламус имеет крайне важную и незаменимую функцию для организма. Трудно представить, но этот сравнительно маленький орган несет ответственность за все психические функции: восприятие и понимание, память и мышление, ведь благодаря ему мы видим, понимаем, ощущаем мир и воспринимаем все, что нас окружает. Благодаря его работе мы ориентируемся в пространстве и во времени, чувствуем боль, этот «коллектор чувствительности» воспринимаем и перерабатывает информацию, полученную от всех рецепторов, кроме обоняния и передает необходимый сигнал в нужный отдел коры головного мозга. В итоге организм дает правильную реакцию, проявляет правильные модели поведения на соответствующий раздражитель или сигнал.
Общие сведения
Промежуточный мозг расположен под мозолистым телом и состоит из: таламуса (таламического мозга) и гипоталамуса.
Таламус (он же: зрительный бугор, коллектор чувствительности, информатор организма) – это отдел промежуточного мозга, находящийся в его верхней части, над стволом мозга. Сюда стекаются сенсорные сигналы, импульсы из самых разных частей организма и от всех рецепторов (кроме обоняния). Тут они перерабатываются, орган оценивает, насколько важны приходящие импульсы для человека и отправляет информацию дальше в ЦНС (центральная нервная система) или к коре головного мозга. Этот кропотливый и жизненно важный процесс происходит благодаря составляющим таламуса – 120 разнофункциональным ядрам, которые несут ответственность за принятие сигналов, импульсов и за отправку переработанной информации в соответствующий .
Благодаря сложной структуре, «зрительный бугор» способен не только принимать и перерабатывать сигналы, но и анализировать их.
Готовая информация о состоянии организма и его проблемах поступает к коре головного мозга, которая, в свою очередь, разрабатывает стратегию решения и устранения проблемы, стратегию дальнейших действий и поведения.
Строение
Таламус - парное яйцевидное образование, состоящее из нервных клеток, которые объединяются в ядра, благодаря которым и происходит восприятия и обработка сигналов и импульсов, идущих от разных органов чувств. Таламус занимает основную часть промежуточного мозга (приблизительно 80%). Состоит из 120 разнофункциональных ядер серого вещества. По форме он напоминает небольшое куриное яйцо.
Исходя из строения и расположения отдельных частей, таламический мозг можно разделить на: метаталамус, эпиталамус и субталамус.
Метаталамус (подкорковый слуховой и зрительный центр) - состоит из медиальных и латеральных коленчатых тел. В ядро медиального коленчатого тела заканчивается слуховая петля, а в латеральную – зрительные тракты.
Медиальные коленчатые тела составляют слуховой центр. В медиальной части метаталамуса из подкоркового слухового центра аксоны клеток направляются к корковому концу слухового анализатора (верхняя височная извилина). Дисфункция этой части метаталамуса может привести к снижению слуха или к глухоте.
Латеральные коленчатые тела составляют подкорковый зрительный центр. Тут заканчиваются зрительные тракты. Аксоны клеток, формируют зрительную лучистость, по которой зрительные импульсы достигают коркового конца зрительного анализатора (затылочная доля). Дисфункция этого центра может привести к проблемам со зрением, а серьезные поражения – к слепоте.
Эпиталамус (надталамус) – верхняя задняя часть таламуса, которая возвышается над ним: включает эпифиз, который является надмозговой железой внутренней секреции (шишковидное тело). Эпифиз находится в подвешенном состоянии, так как расположен на поводках. Он отвечает за выработку гормонов: днем он вырабатывает гормон серотонин (гормон радости), а ночью – мелатонин (регулятор режима дня и гормон ответственный за цвет кожи и глаз). Эпиталамус играет роль в регуляции жизненных циклов, регулирует период наступления полового созревания, режимы сна и бодрствования, тормозит процессы старения.
Поражения эпиталамуса приводят к нарушению жизненных циклов, в том числе к бессоннице, а также к половым дисфункциям.
Субталамус (подталамус) или преталамус является мозговым веществом маленького объема. Состоит в основном из субталамического ядра и имеет соединения с бледным шаром. Субталамус контролирует мышечные ответы и отвечает за выбор действия. Поражение субталамуса приводят к двигательным нарушениям, тремору, параличу.
Кроме всего перечисленного, таламус имеет связи со спинным мозгом, с гипоталамусом, подкорковыми ядрами и, естественно, с корой головного мозга.
Каждый отдел этого уникального органа несет определенную функцию и отвечает за жизненно важные процессы, без которых нормальное функционирование организма невозможно.

Функции таламуса
«Коллектор чувствительности» получает, фильтрует, перерабатывает, интегрирует и направляет в мозг информацию, которая поступает от всех рецепторов (кроме обоняния). Можно сказать, что в его центрах происходит формирование восприятия, ощущения, понимания, после чего обработанная информация или сигнал поступают в кору больших полушарий.
Главными функциями органа являются:
- переработка информации получаемой от всех органов (рецепторы зрения, слуха, вкуса и осязания) чувств (кроме обоняния);
- управление эмоциональными реакциями;
- регулирование непроизвольной двигательной активности и мышечного тонуса;
- поддерживание определенного уровня активности и возбудимости головного мозга, что необходимо для восприятия информации, сигналов, импульсов и раздражений исходящих извне, из окружающей среды;
- отвечает за интенсивность и чувство боли.
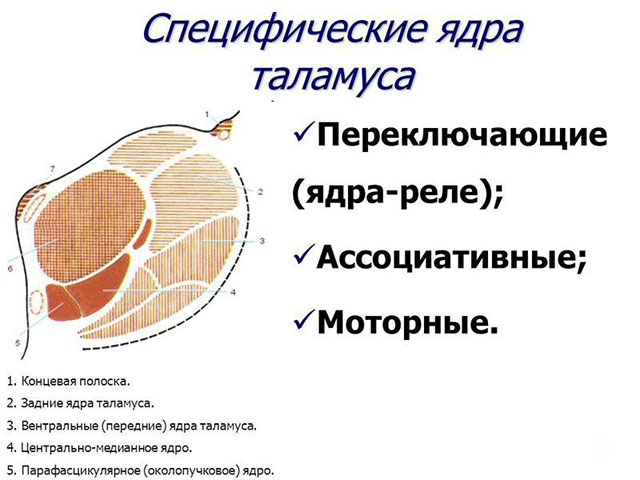
Как мы уже говорили, каждая доля таламуса состоит из 120 ядер, которые исходя из функциональности, можно разделить на 4 основные группы:
- латеральную (боковые);
- медиальную (срединные);
- ассоциативную.
Ретикулярная группа ядер (отвечает за равновесие) – отвечает за обеспечение равновесия при ходьбе и баланса в организме.
Латеральная группа (центр зрения) – отвечает за зрительное восприятие, принимает и передает импульсы в теменную, затылочную часть коры головного мозга – зрительной зоне.
Медиальная группа (центр слуха) - отвечает за слуховое восприятие, принимает и передает импульсы в височную часть коры — слуховой зоне.
Ассоциативная группа (тактильные ощущения) - принимает и передает в кору головного мозга тактильную информацию, то есть сигналы, исходящие от рецепторов кожных покровов и слизистых оболочек: болевые ощущения, зуд, удар, прикосновение, раздражение и т.д.
Также, с функциональной точки зрения, ядра можно разделить на: специфические и неспецифические.
К специфическим ядрам поступают сигналы от всех рецепторов (кроме обоняния). Они обеспечивают эмоциональную реакцию человека и отвечают за возникновение болевых ощущений.
Специфические ядра, в свою очередь, бывают:
- внешние - получают импульсы от соответствующих рецепторов и отправляют информацию в конкретные зоны коры. Благодаря этим импульсам возникают чувства и ощущения;
- внутренние - не имеют прямых связей с рецепторами. Получают информацию уже переработанной со стороны релейных ядер. От них импульсы идут в кору головного мозга в ассоциативные зоны. Благодаря этим импульсам возникают примитивные ощущения и обеспечивается взаимосвязь между сенсорными зонами и корой больших полушарий.
Неспецифические ядра поддерживают общую активность коры головного мозга, посылая неконкретные импульсы и стимулируя мозговую активность. Не имея прямой связи с корой, неспецифические ядра таламуса передают свои сигналы в подкорковые структуры.
Отдельно о зрительном бугре
Ранее считалось, что таламус обрабатывает только зрительные импульсы, тогда орган и получил название — зрительные бугры. Сейчас это название считается устаревшим, так как орган обрабатывает практически весь спектр афферентных систем (кроме обоняния).
Система, которая обеспечивает зрительное восприятие – одна из самых интересных. Основной внешний орган зрения – глаз – рецептор, который имеет сетчатку и оснащен особенными клетками (колбочки, палочки), которые трансформируют световой пучок и электрический сигнал. Электрический сигнал, в свою очередь, проходя по нервным клеткам, попадает в латеральный центр таламуса, который отправляет обработанный сигнал в центральный отдел коры головного мозга. Тут происходит окончательный анализ сигнала, благодаря чему формируется увиденное, то есть – картинка.
Чем опасны дисфункции зон таламуса
У таламуса сложная и налаженная структура, поэтому, если возникают сбои или проблемы в работе даже отдельно взятой зоны органа – это приводит к разным последствиям, влияя на отдельные функции организма и даже на весь организм в целом.
Прежде чем попасть в соответствующий центр коры, сигналы с рецепторов поступают в таламус, а точнее, в его определенную часть. Если определенные ядра таламуса повреждены, то импульс не обрабатывается, не доходит до коры или доходит в необработанной форме, следовательно, кора головного мозга и весь организм не получают нужную информацию.
Клинические проявления дисфункций таламуса зависят от конкретной зоны поражения и могут проявляться: проблемами с памятью, вниманием, пониманием, потерей ориентации в пространстве и во времени, нарушениями двигательной системы, проблемами со зрением, слухом, бессонницей, психическими расстройствами.
Одним из проявлений дисфункций органа может быть специфическая амнезия, которая ведет к частичной потере памяти. В этом случае, человек забывает события, произошедшие после повреждения или поражения соответствующей зоны органа.
Еще одно редкое заболевание, затрагивающее таламус – фатальная бессонница, которая может распространяться на нескольких представителей одной семьи. Болезнь возникает по причине мутации соответствующей зоны таламуса, которая отвечает за регулирование процессов сна и бодрствования. Из-за мутации происходит сбой в правильной работе соответствующего участка, и человек перестает спать.
Таламус – также является центр болевой чувствительности. При поражении соответствующих ядер таламуса возникает невыносимая боль либо, наоборот, полная потеря чувствительности.
Таламус, да и мозг в целом продолжают оставаться не до конца изученными структурами. И дальнейшие исследования сулят большие научные открытия и помощь в познании этого жизненно важного и сложного органа.
ТАЛАМУС [thalamus (PNA, JNA, BNA); син. зрительный бугор ] - парное образование промежуточного мозга, являющееся основным коллектором соматосенсорной информации, идущей через мозговой ствол к коре полушарий головного мозга. Т. правого полушария отделен от Т. левого полушария третьим желудочком на всем протяжении, за исключением места межталамического сращения (adhesio interthalamica, massa intermedia), в к-ром располагаются непарные ядра так наз. средней линии.
На основании эмбриол. исследований В. Гис (1904) впервые выделил основные структурные образования промежуточного мозга: эпиталамус (см. Промежуточный мозг), дорсальный (задний) таламус, вентральный (передний) таламус и гипоталамус (см.). В BNA был выделен также и метаталамус, в состав к-рого входят медиальные и латеральные коленчатые тела. Дальнейшие филоонтогенетические исследования строения и связей промежуточного мозга показали принадлежность дорсальной части ядра латерального коленчатого тела к дорсальному Т., а вентрального ядра латерального коленчатого тела и вентральной крупноклеточной части медиального коленчатого тела - к вентральному Т.
Сравнительная анатомия
Сравнительно-анатомические и эмбриологические исследования выявили весьма сложный характер развития Т. и отдельных его комплексов, определяемый как общими закономерностями эволюции мозга хордовых, так и экологическими факторами формирования сенсорных систем у различных представителей этого типа.
Образования вентрального Т., особенно наиболее древняя вентральная часть латерального коленчатого тела, дифференцируются уже у селахий, т. е. значительно раньше структур дорсального Т., подразделение к-рого на отдельные образования выявляется лишь у рептилий. У птиц и рептилий наиболее развитой структурой дорсального Т. является круглое ядро, не имеющее прямых гомологов в дорсальном Т. млекопитающих. Наиболее сложно дифференцированы структуры дорсального Т. у млекопитающих. Древнейшие образования дорсального Т.- заднее и претектальное ядра, хорошо развитые у низших млекопитающих, у высших - редуцируются. Подобный процесс происходит и с интраламинарными (внутрипластинча-тыми, Т.) ядрами, а также с вентральной частью ядра латерального коленчатого тела, относящимся к вентральному Т.
Усложнение дифференцировки структур дорсального Т. связано с еще большим усложнением строения новой коры (нео-кортекса) мозга млекопитающих, где закономерности дивергентного (разнонаправленного) развития проявляются в относительном прогрессировании новейших и относительном регрессировании филогенетически древнейших зон коры полушарий головного мозга. Этот процесс объясняется тем, что по мере эволюционного усложнения мозга в корково-подкорковых отношениях корково-таламические связи становятся определяющими, а в них, в свою очередь, преобладающими (по сравнению с проекционными, релейными) становятся ассоциативные связи. Так, у обезьян проекционные зоны занимают еще значительно большую территорию коры головного мозга по сравнению с ассоциативными зонами; напр., только затылочная проекционная зона составляет ок. 20% площади всей коры. У человека соотношения обратные - доминирующими являются ассоциативные зоны, и затылочная проекционная зона составляет всего 12% площади новой коры. Поэтому и в таламических комплексах мозга человека наиболее развиты ассоциативные ядра.
Анатомия и микроскопическое строение
Т. четко отграничен от окружающих структур. Гипоталами-ческая борозда, идущая вдоль стенки третьего желудочка от межже-лудочкового отверстия к отверстию сильвиева водопровода (водопровода мозга, Т.), ограничивает Т. от вент-рально расположенного гипоталамуса. На дорсальной поверхности Т. расположена так наз. прикрепленная пластинка, образующая центральную часть дна бокового желудочка мозга; в дорсолатеральном направлении Т. отделен пограничной полоской от структур хвостатого ядра; латерально внутренняя капсула отделяет Т. от чечевицеобразного ядра и головки хвостатого ядра (см. Базальные ядра). Ростральный конец Т. представлен передним бугорком, а каудальный образует подушку, под задним концом к-рой расположены латеральное коленчатое тело, а кнутри от него - медиальное коленчатое тело, входящие в таламический комплекс.
Морфол. классификации ядерных образований Т. имеют в основе топографический принцип и выделяют переднюю, заднюю, наружную и внутреннюю группы ядер. Развитие современных комплексных морфо-физиол. методов исследования обусловило возможность выявления и структурно-функциональной дифференцировки таламических ядер, к-рые подразделяют на: 1) специфические сенсорные (релейные, переключающие) ядра, в каждое из к-рых поступает с периферии преимущественно какой-либо один основной вид сенсорной афферентации; 2) неспецифические ядра, принимающие и перерабатывающие импульсы, поступающие прежде всего из ретикулярных образований мозгового ствола и гипоталамуса; 3) ассоциативные ядра, имеющие отношение к дальнейшей переработке и интеграции различных сенсорных импульсов. Выделение ассоциативной группы ядер связано с новыми данными об интегрирующих (а не только о релейных) функциях таламических структур.
Передняя группа ядер, находящаяся в переднем бугорке Т., включает переднедорсальное и передневентральное ядра, относящиеся к специфическим релейным образованиям, и переднемедиальное ядро, относящееся к неспецифическим структурам (рис., а). Релейные ядра получают импульсы по сосцевидно-таламическому пучку и передают их в пре- и постсубикулярные поля старой коры, но гл. обр.- в поля лимбической области новой коры. Эти образования входят в состав лимбической системы (см.). Задняя группа ядер заключена в заднем полюсе Т. (подушке) и является ассоциативным ядром, наиболее развитым у приматов, и особенно у человека. Структурно и функционально задняя группа имеет отношение не только к зрительным (в большей мере), но и к соматосенсорным и слуховым (в меньшей мере) проекциям на кору полушарий головного мозга.
В наружный комплекс ядер (вент-ролатеральные ядра, Т.) входят латеральное переднее (вентральное) ядро и вентральная группа ядер, в к-рой выделяют передние и задние вентральные ядра. Латеральное переднее ядро - филогенетически наиболее древнее образование комплекса, расположено кзади от передней группы ядер и четко отграничено от окружающих структур; имеет прямые связи со сводом мозга, что позволяет отнести ядро к релейным образованиям лимбической системы (рис., б). Заднее латеральное ядро относится к ассоциативным ядрам Т., однако предполагается, что это ядро имеет отношение и к специфическим сенсорным системам.
Основным релейным ядром Т., передающим соматическую афферен-тацию в кору головного мозга, является заднемедиальное вентральное, или постеромедиальное ядро, к к-ро-му подходит медиальная петля, спинно-таламический путь, мозжечково-рубро-таламический путь и аксоны клеток ядра спинномозгового пути тройничного нерва.
Внутренняя группа ядер (медиальные ядра, Т.) таламуса располагается снаружи от центрального серого вещества, окружающего стенки третьего желудочка (см. Желудочки головного мозга). Наиболее развитым у приматов и человека является дорсальное медиальное ядро, относящееся к ассоциативным ядрам и имеющее прямые связи со структурами лобной области новой коры. Внутри пластинки, окружающей это ядро, располагаются интраламинар-ные (внутрипластинчатые) ядра. К этой же группе относят и парафас-цикулярный комплекс ядер, кнаружи от к-рого располагается срединный центр Люиса (centrum medianum Luysi) или центральное срединное ядро, вопрос о прямых связях к-рого с корой полушарий головного мозга до наст, времени недостаточно изучен. Медиальное вентральное ядро, расположенное под дорсальным медиальным, является филогенетически более древним образованием и может быть отнесено к неспецифическим структурам так же, как и ядра средней линии, расположенные внутри центрального серого вещества.
Еще к более древним структурам Т. принадлежат претектальное и заднее ядра.
Латеральное коленчатое тело, относящееся к таламическому комплексу состоит из филогенетически более новой дорсальной части ядра (дорсальное ядро) и более старой вентральной части (вентральное ядро). Дорсальное ядро является классическим релейным образованием, передающим зрительную аф-ферентацию в кору (см. Зрительный анализатор). Терминали от сетчатки глаз находят и в вентральной части ядра латерального коленчатого тела, а волокна от затылочной области коры распределяются как в вентральной, так и в дорсальной частях ядра латерального коленчатого тела.
Медиальное коленчатое тело, особенно его наружная мелкоклеточная часть, является основным релейным образованием слухового анализатора (см.), принимающим информацию по волокнам латеральной петли и передающим ее в структуры височной области новой коры.
Кровоснабжение Т. - см. Мозговое кровообращение.
Физиология
Исследование структуры и функции таламических образований с помощью современных комплексных методов выявило чрезвычайно сложньш характер отношений Т. как с выше расположенными отделами переднего мозга, так и с находящимися ниже структурами мозгового ствола, мозжечка и спинного мозга. Тесная интеграция соматических, висцеральных, мотивационных влияний, осуществляемая на уровне таламических комплексов, определяет ведущую роль этих образований промежуточного мозга в формировании не только корковоподкорковых отношений, но и в становлении и развитии целостных поведенческих актов животных и человека.
Изучение функций Т. проводят путем его электрического раздражения или разрушения в сочетании с анализом динамики различных поведенческих реакций, регистрацией ЭЭГ, вызванных потенциалов или электрической активности одиночных нейронов в ответ на различные периферические раздражения в экспериментах на животных и в леч. целях - у человека.
Установлено, что в релейных ядрах Т. переключаются импульсы тактильной, кинестетической, температурной, болевой (локализованная боль), вкусовой и висцеральной чувствительности (см.). В каждое ядро поступают импульсы с противоположной стороны тела, лишь область лица имеет в заднем вентральном ядре Т. билатеральное представительство. Части тела, выполняющие наиболее интенсивные сенсорно-двигательные функции (лицо, язык, дистальные отделы конечностей), имеют более обширное представительство. В медиальный отдел заднего вентрального ядра, сомато-топическая организация к-рого хорошо изучена, приходит афферента-ция от ростральных частей тела и импульсы от вкусовых рецепторов, в латеральный отдел - от рецепторов туловища и конечностей. Проекции тактильной и висцеральной чувствительности имеют в Т. широкое перекрытие, что может обусловливать феномен иррадиации висцеральной боли (см.) на поверхности тела. Общие проекции в Т. имеют также тактильная и глубокая (про-приоцептивная) чувствительность. Тонкодифференцированная афферентная импульсация от термо- и ноцирецепторов проходит преимущественно по спиноталамическому пути в заднее вентральное ядро, тогда как более грубые формы этих видов чувствительности (диффузная боль, дифференцировка больших температурных диапазонов) связаны преимущественно с неспецифическими интраламинарными ядрами Т. (парафасцикулярным комплексом), а также, возможно, с задними ядрами. Электростимуляция заднего вентрального ядра у человека в процессе стереотаксических операций (см. С тереотаксическая нейрохирургия, Стереотаксический метод) вызывает парестезии, реже нарушение схемы тела (см. Схема тела).
К несенсорным релейным образованиям относят переднюю группу ядер, передающую импульсацию от сосцевидных тел к лимбической области коры, а также два ядра вентральной группы Т. - переднее вентральное и латеральное вентральное, передающие основную массу возбуждений из мозжечка и бледного шара к моторным отделам коры головного мозга, что и определяет их роль в регуляции непроизвольной двигательной активности и мышечного тонуса. Указанные связи латерального вентрального ядра обосновывают целесообразность его деструкции у больных паркинсонизмом и с различными гиперки-незами.
Группа ассоциативных ядер Т. имеет основным источником аффе-рентации релейные и отчасти неспецифические структуры Т. и гипоталамус. В последнее время показано, что в этих ядрах может заканчиваться часть сенсорных проекций (соматосенсорных, слуховых, зрительных, висцеральных). Взаимодействие различных афферентных импульсов на нейронах ассоциативных ядер Т., их сложные внутрита-ламические и межталамические связи объясняют важную роль этой группы образований в интегративной деятельности мозга. Различают таламопариетальную и таламофрон-тальную ассоциативные системы. Первая имеет преимущественное отношение к формированию комплексов афферентных воздействий, необходимых для оценки сигнальной значимости сложных видов раздражителей, лежащей в основе гностической функции, особенно зрительного и слухового восприятия, а также сенсорного обеспечения управления произвольными движениями. Медиальное дорсальное ядро принадлежит таламофронтальной ассоциативной системе, участвующей прежде всего в аффективно-эмоциональной и программирующей деятельности. Стимуляция медиального дорсального ядра Т. у больных вызывает разнообразные положительные и отрицательные эмоциональные реакции; деструкция этого ядра в леч. целях, а также в экспериментах на животных ослабляет чувство страха, тревоги и напряженности, но при этом возникают нерезко выраженные симптомы так наз. лобного синдрома - снижение инициативы и интеллектуальных способностей на фоне стойкого уменьшения эмоциональной реактивности. При распространении патол. процесса на медиальные отделы Т. наблюдают развитие слабоумия (см.).
Группа неспецифических ядер Т. получает афферентацию прежде всего из ретикулярных образований мозгового ствола, однако есть данные о возможности окончания в hYix волокон, идущих в составе специфических восходящих систем. Хотя ядра этой группы проецируются на кору более диффузно, чем первые две группы, нек-рым из них свойствен более или менее локальный характер связей с корой и полосатым телом (стриопаллидумом). К этой группе относят внутрипластинчатые (интраламинарные) ядра (парафасци-кулярное ядро, латеральное центральное и парацентральное ядра), ретикулярное ядро таламуса, ядра средней линии Т. Открытие неспеци-фицеских ядер Т. связано с работами Демпси и Морисон (E. W. Dempsey, R. S. Morison, 1943), показавшими, что при раздражении этих ядер в коре головного мозга возникает распространенная реакция вовлечения. Позднее С. П. Нарикашвили, Э. С. Мониа-ва, В. И. Гусельников и А. Я. Супин (1968) и др. обнаружили, что в генезе этой реакции, так же как и реакции усиления, ведущую роль играет взаимодействие несиецифических ядер Т. со специфическими его образованиями.
Данные о функциональном значении ядер этой группы разноречивы: с одной стороны, установлено, что разрушение комплекса парафасцику-лярное ядро - центральное срединное ядро (срединный центр) влияет на позитивное и негативное обучение крыс; с другой стороны, эффект повреждения этого комплекса в значительной степени зависит от предшествующего эмоционального фона; показана также важная роль афферентации данного комплекса в деятельности нейронов хвостатого ядра (см. Базальные ядра). Представление о неспецифичности этой группы ядер относительно. На основании данных об их строении, связях и функции можно говорить о существовании в Т. сложных комплексов, образованных релейными и неспецифическими ядрами. Изучение функций ядер средней линии затруднено их теснейшей близостью к ассоциативным и релейным ядрам Т. Разрушение медиальных отделов Т. может сопровождаться нарушениями условных рефлексов (см.), памяти (см.), сна (см.).
В группе неспецифических ядер особое место занимает ретикулярное ядро Т. Последнее примыкает снаружи к различным релейным ядрам дорсального Т. и частично имеет общую с ними проекцию на кору. Предполагают, что это ядро является посредником как в деятельности других ядер Т., так и в таламо-кортикальном взаимодействии его специфических и неспецифических образований. В наст, время очевидна важная роль Т. в осуществлении сложных функций восприятия и переработки различных сигналов, поступающих далее в кору головного мозга и базальные ядра, в поддержании уровня бодрствования и эмоционального состояния, в нормальном обеспечении простых и более сложных форм поведения и памяти. Полное двустороннее удаление Т. у собак и кошек обнаруживает резкий дефицит приспособительных реакций, нарушения чувствительности и чередований цикла бодрствование - сон. Оперативная изоляция коры и базальных ядер от Т. и мозгового ствола (так наз. таламическое животное) приводит к массивной ретроградной дегенерации нейронов Т. и сопровождается резкими и необратимыми изменениями адаптивного поведения, вплоть до невозможности самостоятельного питания.
В последнее время в связи с накоплением данных о межполушар-ной асимметрии появились указания на функциональную неравнозначность правого и левого Т. у человека. Так, использование вербализуемого материала в тестах на кратковременную память, видимо, осуществляется с участием преимущественно левого Т., невербализуемого материала - с участием правого Т.; наиболее существенным синдромом левостороннего поражения таламуса является несодержательность речи.
В 70-х гг. был сформулирован ряд новых принципов таламокортикаль-ного взаимодействия, имеющих отношение к пониманию интеграции нервных процессов на уровне переднего мозга: принципы «ядерного» и «рассеянного» типов таламокорти-кальных проекций, переслаивания этих различных проекций между собой, их различного распределения в проекционных и ассоциативных областях новой коры, моно- и оли-госинаптической передачи возбуждений из Т. в кору, а также топографического соответствия (преимущественно для релейных и ассоциативных структур) и несоответствия (гл. обр. для неспецифических структур Т.) таламокортикальных и кор-тико-таламических влияний. В ней-рофизиол. исследованиях обнаружена динамическая интеграция специфических и неспецнфических влияний на диэнцефальном и корковом уровнях, установлено, что ритмическая активность коры головного мозга обусловлена активностью таламических ядер, что формирование a-ритма в коре происходит при участии как таламических, так и корковых структур. Обнаружено, что нейрофизиол. основой ритмической активности головного мозга являются механизмы возвратного торможения (см.) п возбуждения (см.), разыгрывающиеся на таламическом и корковом уровнях. Выделены и изучены новые пути взаимоотношений ассоциативных и проекционных таламокортикальных систем и сформулировано положение о сложно функционирующих таламокортико-таламических ассоциативных системах. Получила новое освещение проблема корковой регуляции деятельности таламических ядер: показано, что регуляция релейных таламических ядер осуществляется гл. обр. прямыми кортико-таламиче-скими путями, а неспецифических ядер - преимущественно с участием ретикулярной формации мозгового ствола. Установлено, что ретикулярные системы мозга способны дифференцировать афферентные сигналы и направлять их в соответствующие структуры мозга под влиянием кортико-фугальной модуляции.
Патология
Очаговые поражения Т. могут развиваться при различных патол. процессах, однако чаще всего - при нарушениях мозгового кровообращения (ишемия, кровоизлияние на почве атеросклероза и гипертонической болезни). Наиболее часты нарушения кровообращения в зоне глубоких ветвей задней мозговой артерии. При инсульте в этом бассейне в основном повреждаются вен-тролатеральные ядра Т. или его дорсальное медиальное ядро. В Т. возможны как ишемические инфаркты, так и кровоизлияния при разрывах артерий, васкулитах, гипертонических кризах с высоким АД, при артериовенозных аневризмах. Значительно реже таламические комплексы повреждаются при инфекционных, опухолевых, дистрофических и других заболеваниях.
Воспалительные поражения Т. изредка встречаются при туберкулезе, сифилисе, сепсисе, при энцефалитах (вирусных, бактериальных). Дегенеративные процессы в структурах Т. возникают при генетических дефектах обмена веществ или при интоксикациях (экзогенных, эндогенных), а также в резидуальной стадии инфекционного или травматического поражения мозга. Первичные опухоли Т. сравнительно редки, чаще Т. вовлекается в патол. процесс при инфильтративном росте опухоли из соседних отделов мозга (при глиомах, астроцитомах и др.).
Клин, симптомы при поражении Т. многообразны и зависят от функциональной роли поврежденных структур. Таламический синдром впервые подробно описан Ж. Деже-рином и Г. Русси в 1906 г. при размягчении Т. вследствие закупорки задней мозговой артерии и ее ветвей. При выключении a. thalamoge-niculata на стороне, противоположной очагу поражения в Т., развиваются следующие симптомы: 1) ге-мигипестезия или гемианестезия с выраженным нарушением глубокой чувствительности больше на конечностях, иногда без расстройства чувствительности на лице; 2) гиперпатия или дизестезии (см. Чувствительность, расстройства), пароксизма л ьно возникающие или постоянные жестокие боли, распространяющиеся на всю половину тела (таламический болевой синдром); 3) утрата вибрационной чувствительности; 4) преходящий гемипарез без выраженной мышечной спастичности и патол. рефлекса Бабинского; 5) атрофия мышц пострадавшей половины тела; 6) хореические и атетоид-ные движения в пальцах руки; 7) гемиатаксия; 8) иногда гомонимная гемианопсия; 9) нотнагелевский мимический парез; 10) расстройства внимания.
При частичном поражении Т. могут преобладать отдельные группы этих симптомов. При разрушении медиальной части Т. (бассейн a. tha-lamoperforata) повреждается дентато-руброталамический путь, по к-рому в Т. поступают импульсы из мозжечка, при этом возникает гиперкинез (атетоидный, хореический) и гемиатаксия на противоположной очагу стороне. При поражении передних отделов Т. выявляют контралатеральный мимический парез лицевой мускулатуры. При диффузном поражении Т. таламический синдром нередко сочетается с вегетативными нарушениями (цианоз, атрофия кожи и ногтей, пастозность, похолодание кожи на пораженной стороне тела), более выраженными в руке. При этом рука приобретает характерное положение: предплечье согнуто и прони-ровано, кисть согнута в лучезапястном суставе, проксимальные фаланги пальцев согнуты, в то время как средние и дистальные фаланги разогнуты (так наз. таламическая рука). Иногда на стороне очага возникает синдром Бернара - Горнера (см. Бернара - Горнера синдром).
Диагностика таламического синдрома основывается на выявлении комплекса составляющих его симптомов. Клин, методы диагностики состоят в определении поверхностной и глубокой чувствительности, определении полей зрения (см.), силы мышц конечностей, координации движений (см.) и др. Диагностике помогает симптом Ферстера: раздражение различных рецепторов (зрительных, слуховых, вкусовых) вызывает боль и неприятное ощущение в пострадавшей половине тела.
Дифференциальный диагноз таламического синдрома Дежерина - Русси следует проводить с синдромом Геда - Холмса - односторонней аффективной дизестезией, для к-рого характерны односторонняя дизесте-зия, утрированные внешние проявления аффективных реакций (гримасничание), резкие защитные движения в ответ на незначительное физическое раздражение (напр., легкий укол булавкой), одностороннее расстройство вкуса и обоняния. Этот симптомокомплекс развивается при нарушении корково-таламических и корково-гипоталамических связей. Кроме того, таламический синдром следует дифференцировать с синдромом поражения внутренней капсулы, при к-ром наряду с гемианестезией (иногда в сочетании с гемианопсией) наблюдается спастическая гемиплегия. Поражение Т. можно выявить при церебральной ангиографии (см.), компьютерной томографии (см. Томография компьютерная).
Лечение таламического синдрома патогенетическое. Значительные трудности возникают при таламической гемиалгии, при к-рой обычные анальгетики мало эффективны. Рекомендуют применять седуксен, аминазин, финлепсин, тегретол и др. При гиперкинезах и неустранимых болевых синдромах применяют стереотаксические операции на ядрах Т. (см. Стереотаксическая нейрохирургия).
Прогноз таламического синдрома зависит от характера основного патол. процесса.
Библиография: Адрианов О. С. О принципах организации интегративной деятельности мозга, М., 1976; Батуев А. С. Высшие интегративные системы мозга, Д., 1981, библиогр.; ГусельниковВ. И. Электрофизиология головного мозга, М., 1976; Дуринян Р. А. Центральная структура афферентных систем, Л., 1965; он же, Корковый контроль неспецифических систем мозга, М., 1975; Кесарев В. С. Количественная архитектоника мозга человека, Вестн. АМН СССР, № 12, с. 29, 1978; Клиническая нейрофизиология, под ред. Н. П. Бехтерева, с. 49, Л., 1972; Кроль М. Б. и Федорова Е. А. Основные невропатологические синдромы, М., 1966; Курепина М. М. Мозг животных, М., 1981;Общая и частная физиология нервной системы, под ред. П. Г. Костюка, с. 313, Л., 1969; Серков Ф. Н. и Казаков В. Н. Нейрофизиология таламуса, Киев, 1980, библиогр.; Сосудистые заболевания нервной системы, под ред. Е. В. Шмидта, с. 56, 330, М., 1975; Экспериментальная психология, под ред. С. С. Стивенса, пер. с англ., с. 174, М., 1960; В a r r a q u е r-В о г- d a s L. а. о. Thalamic hemorrhage, Stroke, v. 12, p. 524, 1981; Brow n J. W. Thalamic mechanisms in language, Handb. behavioral neurobiol., ed. by F. A. King, v. 2, p. 215, N. Y, -L., 1979; DantzerR. etDela-c о u r J. Modification d’un phenomene de suppression conditionnee par une leson thalamique, Physiol. Behav., t. 8, p. 997, 1972; Dejerine J. a. Roussy G. Le syndrome thalamique, Rev. neurol. (Paris), t. 14, p. 521, 1906; Einfiihrung in die stereotaktischen Operationen mit einem Atlas des menschlichen Gehirns, hrsg. v. G. Schaltenbrand a. P. Bailey, Bd 1, S. 230, 1959, Bibliogr.; Graeber R. C. a. Ebbesson S.O. Retinal projections in the lemon shark (Negaprion previrostris), Brain Behav. Evolut., v. 5, p. 461, 1972; G r a y b i e 1 A. M. Some fiber pathways related to the posterior thalamic region in the cat, ibid., v. 6, p. 363, 1972; Henderson V. W., Alexander M. P. a. N a e s e г M. A. Right thalamic injury, impaired visuospatial perception, and alexia, Neurology, v. 32, p. 235, 1982, bibliogr.; K w a k R., K a d о у a S. a. Suzuki T. Factors affecting the prognosis in thalamic hemorrhage, Stroke, v. 14, p. 493, 1983; Lapresle J. a. Ha-g и e n а и M. Anatomico-chemical correlation in focal thalamic lesions, Z. Neurol., v. 205, p. 29, 1973; L e j e u n e H. Lesions thalamiques medianes et effets differentiels dans les apprentissages operants, Physiol, a. Behav., t. 18, p. 349, 1977; Van В u-r e n J. М. а. В o r k e R. C. Variations and connections of the human thalamus, pt 1 - 2, N. Y. a. o., 1972; V e 1 a s с o F. a. Velasco M. A reticulothalamic system mediating proprioceptive attention and tremor in man, Neurosurgery, v. 4, p. 30, 1979; Villablanca J. a. Sali-nas-Zeballos М. E. Sleep-wakeful-ness, EEG and behavioral studies of chronic cats without the thalamus, Arch. ital. Biol., v. 110, p. 383, 1972.
Д. К. Богородинский, А. А. Скоромец; О. С. Адрианов (физ.), В. С. Кесарев (ан., сравнительная анатомия).
Таламус – зрительный бугор
Структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка.
В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути. Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор совместно с гипоталамусом принимает участие в формировании и реализации инстинктов, влечений, эмоций. В таламусе около 120 разнофункциональных ядер, которые образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная - в лобную долю коры; латеральная - в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая - в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральное, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно. Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов. От специфических ядер импульсы поступают в строго определенные участки III-IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую организацию. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т.е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность, т.е. способность реагировать на определенную частоту звуковых колебаний. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное - с лобной долей коры, латеральное дорсальное - с теменной, подушка - с ассоциативными зонами, теменной и височной долями коры большого мозга.
На полисенсорных нейронах таламуса происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра - с теменной, нейроны медиального ядра - с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены: срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т.е. развитие сонного состояния.
Гипоталамус (подбугровая область) – изложение материала будет в разделе «Лимбическая система».
Мозжечок
Мозжечок (cerebellum) -структура головного мозга, принимающая участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций. Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка:
1) кора мозжечка построена достаточно однотипно, имеет стереотипные связи, что создает условия для быстрой обработки информации;
2) основной нейронный элемент коры - клетка Пуркинье, имеет большое количество входов и формирует единственный аксонный выход из мозжечка, коллатерали которого заканчиваются на ядерных его структурах;
3) на клетки Пуркинье проецируются практически все виды сенсорных раздражений: проприоцептивные, кожные, зрительные, слуховые, вестибулярные и др.;
4) выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом.
Мозжечок анатомически и функционально делится на старую, древнюю и новую части.
Из мозжечка информация уходит через верхние и нижние ножки. Через верхние ножки сигналы идут в таламус, в мост, красное ядро, ядра ствола мозга, в ретикулярную формацию среднего мозга. Через нижние ножки мозжечка сигналы идут в продолговатый мозг к его вестибулярным ядрам, оливам, ретикулярной формации. Средние ножки мозжечка связывают новый мозжечок с лобной долей мозга.
Импульсная активность нейронов регистрируется в слое клеток Пуркинье и гранулярном слое, причем частота генерации импульсов этих клеток колеблется от 20 до 200 в секунду. Клетки ядер мозжечка генерируют импульсы значительно реже - 1-3 импульса в секунду.
В кору мозжечка от кожных рецепторов, мышц, суставных оболочек, надкостницы сигналы поступают по так называемым спиномозжечковым трактам: по заднему (дорсальному) и переднему (вентральному). Эти пути к мозжечку проходят через нижнюю оливу продолговатого мозга. От клеток олив идут так называемые лазающие волокна, которые ветвятся на дендритах клеток Пуркинье.
Ядра моста посылают афферентные пути в мозжечок, образующие мшистые волокна, которые оканчиваются на клетках-зернах III слоя коры мозжечка. Между мозжечком и синим пятном существует афферентная связь с помощью адренергических волокон. Эти волокна способны диффузно выбрасывать норадреналин в межклеточное пространство коры мозжечка, тем самым гуморально изменяют состояние возбудимости его клеток.
Аксоны клеток III слоя коры мозжечка вызывают торможение клеток Пуркинье и клеток-зерен своего же слоя.
Клетки Пуркинье в свою очередь тормозят активность нейронов ядер мозжечка. Ядра мозжечка имеют высокую тоническую активность и регулируют тонус ряда моторных центров промежуточного, среднего, продолговатого, спинного мозга.
Подкорковая система мозжечка состоит из трех функционально разных ядерных образований: ядра шатра, пробковидного, шаровидного и зубчатого ядра.
Ядро шатра получает информацию от медиальной зоны коры мозжечка и связано с ядром Дейтерса и РФ продолговатого и среднего мозга. Отсюда сигналы идут по ретикулоспинальному пути к мотонейронам спинного мозга.
Промежуточная кора мозжечка проецируется на пробковидное и шаровидное ядра. От них связи идут в средний мозг к красному ядру, далее в спинной мозг по руброспинальному пути. Второй путь от промежуточного ядра идет к таламусу и далее в двигательную зону коры большого мозга.
Зубчатое ядро, получая информацию от латеральной зоны коры мозжечка, связано с таламусом, а через него - с моторной зоной коры большого мозга.
Контроль двигательной активности мозжечком обеспечивается эфферентными сигналами к спинному мозгу, и регулируют силу мышечных сокращений, обеспечивают способность к длительному тоническому сокращению мышц, способность сохранять оптимальный тонус мышц в покое или при движениях, совершать адекватные произвольные движения, быстро переходить от сгибания к разгибанию и наоборот.
Мозжечок обеспечивает координированные сокращения разных мышц при сложных движениях. Например, при ходьбе, когда человек делает шаг, то одновременно центр тяжести туловища переносится вперед. В тех случаях, когда мозжечок не выполняет своей регуляторной функции, у человека наблюдаются расстройства двигательных функций, что выражается следующими симптомами.
1) астения (astenia - слабость) - снижение силы мышечного сокращения, быстрая утомляемость мышц;
2) астазия (astasia, от греч. а - не, stasia - стояние) - утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т.д.;
3) дистония (distonia - нарушение тонуса) - непроизвольное повышение или понижение тонуса мышц;
4) тремор (tremor - дрожание) - дрожание пальцев рук, кистей, головы в покое; этот тремор усиливается при движении;
5) дисметрия (dismetria - нарушение меры) - расстройство равномерности движений, выражающееся либо в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку за предмет (гиперметрия) или не доносит ее до предмета (гипометрия);
6) атаксия (ataksia, от греч. а - отрицание, taksia - порядок) - нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности.
Проявлениями атаксии являются также, асинергия, пьяная шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз-вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Большинство движений человек выучивает в течение жизни, и они становятся автоматическими (ходьба, письмо и т.д.). Когда нарушается функция мозжечка, движения становятся неточными, разбросанными, часто не достигают цели. Данные о том, что повреждение мозжечка ведет к расстройствам приобретенных движений, позволяют сделать вывод, что само обучение шло с участием мозжечковых структур, а, следовательно, мозжечок принимает участие в организации процессов высшей нервной деятельность.
И другие образования .
Таламус расположен латеральнее III желудочка. Он занимает дорсальную часть промежуточного мозга и отделяется от нижележащего бороздой. Два таламуса соединены по средней линии у 70% людей посредством межталамической промежуточной ткани серого вещества. От базальных ядер таламус отделяется внутренней капсулой, состоящей из нервных волокон, соединяющих кору со стволовыми структурами и спинным мозгом. Многие волокна внутренней капсулы продолжают ход в каудальном направлении в составе ножек мозга.
Ядра и функции таламуса
В таламусе выделяют до 120 ядер серого вещества . По месту их расположения ядра делят на передние, латеральные и медиальные группы. В задней части латеральной группы ядер таламуса выделяют подушку, медиальное и латеральное коленчатые тела.
анализ, отбор и передача в кору головного мозга сенсорных сигналов , поступающих к нему из большинства сенсорных систем ЦНС. В этой связи таламус называют воротами, через которые в поступают различные сигналы ЦНС. По выполняемым функциям ядра таламуса делятся на специфические, ассоциативные и неспецифические.
Специфические ядра характеризуются несколькими общими особенностями. Все они получают сигналы от вторых нейронов длинных восходящих афферентных путей, проводящих в кору мозга соматосенсорные, зрительные, слуховые сигналы. Эти ядра, иногда называемые сенсорными, передают обработанные сигналы в хорошо очерченные области коры — соматосенсорную, слуховую, зрительную сенсорные области, а также в премоторную и первичную моторные области коры. С нейронами этих областей коры специфические ядра таламуса имеют реципрокные связи. Нейроны ядер дегенерируют при разрушении (удалении) специфических областей коры, в которые они проецируются. При низкочастотной стимуляции специфических таламических ядер регистрируется усиление активности нейронов в тех областях коры, в которые нейроны ядер посылают сигналы.
К специфическим ядрам таламуса подходят волокна проводящих путей от коры, и ядер ствола мозга. По этим путям могут передаваться как возбуждающие, так и тормозные влияния на активность нейронов ядер. Благодаря таким связям кора мозга может регулировать потоки идущей к ней информации и отбирать наиболее значимую в данный момент. При этом кора может блокировать передачу сигналов одной модальности и облегчать передачу другой.
Среди специфических ядер таламуса имеются также несенсорные ядра. Они обеспечивают обработку и переключение сигналов не от чувствительных восходящих путей, а от других областей мозга. К нейронам таких ядер поступают сигналы от красного ядра, базальных ганглиев, лимбической системы, зубчатого ядра мозжечка, которые после их обработки проводятся к нейронам моторной коры.
Ядра передней группы таламуса участвуют в передаче сигналов от мамиллярных тел к лимбической системе, обеспечивая круговую циркуляцию нервных импульсов по кольцу: лимбическая кора — гиппокамп — — миндалевидное тело — таламус — лимбическая кора. Нейронную сеть, сформированную этими структурами, называют кругом (кольцом) Пайпеца. Циркуляция сигналов по структурам этого круга связана с запоминанием новой информации и формированием эмоций — эмоциональное кольцо Пайпеца.
Ассоциативные ядра таламуса расположены преимущественно медиодорсально, латерально и в ядре подушки. Они отличаются от специфических тем, что к их нейронам не поступают сигналы из чувствительных восходящих путей, а поступают сигналы уже обработанные в других нервных центрах и ядрах таламуса. Ассоциативность нейронов этих ядер выражается в том, что на один и тот же нейрон ядра приходят сигналы разных модальностей. Изменение активности нейронов ядер может быть связано (ассоциировано) с поступлением разнородных сигналов из разных источников (например, от центров, обеспечивающих зрительную, тактильную и болевую чувствительность).
Нейроны ассоциативных ядер являются полисенсорными и обеспечивают возможность осуществления интегративных процессов, в результате которых формируются обобщенные сигналы, передающиеся в ассоциативные области коры лобной, теменной и височной долей мозга. Потоки этих сигналов способствуют осуществлению корой таких психических процессов, как узнавание предметов и явлений, согласование речевых, зрительных и двигательных функций, формирование представления о позе тела, трехмерности пространства и положении в нем тела человека.
Неспецифические ядра таламуса представлены преимущественно интраламинарными, центральными и ретикулярными группами ядер таламуса. Они состоят из мелких нейронов, к которым по многочисленным синаптическим связям поступают сигналы от нейронов других ядер таламуса, лимбической системы, базальных ядер, гипоталамуса, ствола мозга. По чувствительным восходящим путям к неспецифическим ядрам поступает сигнализация от болевых и температурных рецепторов, а по сетям нейронов ретикулярной формации — сигнализация практически от всех других сенсорных систем .
Эфферентные пути от неспецифических ядер идут ко всем зонам коры как непосредственно, так и через другие талами- ческие и ретикулярные ядра. От неспецифических ядер таламуса начинаются также нисходящие пути к стволу мозга. При повышении активности неспецифических ядер таламуса (например, при электрической стимуляции в эксперименте) регистрируется диффузное повышение нейронной активности практически во всех областях коры больших полушарий.
Принято считать, что неспецифические ядра таламуса благодаря своим многочисленным нейронным связям обеспечивают взаимодействие, координацию работы различных областей коры и других отделов головного мозга. Они оказывают модулирующее влияние на состояние активности нервных центров, создают условия для их оптимальной настройки на выполнение работы.
Нейроны различных ядер таламуса оказывают эффекты через высвобождение ГАМК из нервных окончаний, формирующих синапсы на нейронах бледного шара, нейронах локальных цепей, нейронах ретикулярного ядра латерального коленчатого тела; возбуждающие глутамат и аспартат в кортикоталамических, мозжечковых терминалях; таламокортикальных проекционных нейронах. Нейронами секретируются несколько нейропептидов преимущественно в окончаниях восходящих трактов (субстанция Р, сомагостатин, нейропептид Y, энкефалин, холецистокинин).
Метаталамус
Метаталамус включает два таламических ядра — медиальное коленчатое тело (MKT) и латеральное коленчатое тело (ЛКТ).
Ядро медиального коленчатого тела является одним из ядер слуховой системы. Его получают афферентные волокна из латерального лемниска прямо или более часто, после их синаптического переключения на нейронах нижних холмиков. Эти слуховые волокна достигают MKT через соединительную ветвь нижних холмиков. MKT получает также волокна обратной связи из первичной слуховой коры височной области. Эфферентный выход ядра MKT формирует слуховую радиацию внутренней капсулы, волокна которой следуют к нейронам первичной слуховой коры (поля 41, 42).
Нейроны MKT вместе с нейронами нижних холмиков среднего мозга формируют нейронную сеть, выполняющую функцию первичного центра слуха. В нем осуществляется недифференцированное восприятие звуков, их первичный анализ и использование для формирования настораживания, повышения внимания и организации рефлекторного поворота глаз и головы в сторону неожиданного источника звука.
Ядро латерального коленчатого тела является одним из ядер зрительной системы. Его нейроны получают афферентные волокна от ганглиозных клеток обоих сетчаток по зрительному тракту. Ядро ЛКТ представлено нейронами, расположенными в нескольких слоях (пластинках). Сигналы из сетчатки поступают в ЛКТ так, что ипсилатеральная сетчатка проецируется к нейронам 2, 3 и 5-го слоев; контралатеральная — к нейронам 1,4 и 6-го слоев. К нейронам ЛКТ поступают также волокна обратной связи из первичной зрительной коры затылочной доли (поле 17). Нейроны ЛКТ, получив и обработав зрительные сигналы сетчатки, посылают сигналы по эфферентным волокнам, формирующим зрительную радиацию внутренней капсулы в первичную зрительную кору затылочной доли. Некоторые волокна проецируются в ядро подушки и вторичную зрительную кору (поля 18 и 19).
Латеральные коленчатые тела вместе с верхними холмиками относят к подкорковым зрительным центрам. В них осуществляется недифференцированное восприятие света, его первичный анализ и использование для формирования настораживания, повышения внимания и организации рефлекторного поворота глаз и головы в сторону неожиданного источника света.
Внутренняя капсула представляет собой широкий плотный пучок афферентных и эфферентных нервных волокон, соединяющих ствол и кору больших полушарий мозга. Волокна внутренней капсулы продолжаются рострально до радиации мозга и каудально до ножек мозга. Во внутренней капсуле проходят волокна таких важнейших нейронных нисходящих путей, как кортикоспинальный, кортикобульбарный, кортикорубральный, кортикоталамический, лобномостовой, кортикотекальный, кортиконигральный, кортикотегментальный и волокна восходящих таламокоркового, слухового и части зрительного путей.
Во внутренней капсуле тесно располагаются кортикоталамические и таламокортикальные волокна, поэтому при кровоизлияниях и заболеваниях этой области мозга возникают нарушения, характеризующиеся большим разнообразием, чем при повреждении какой-либо другой области ЦНС. Они могут проявиться развитием контралагеральной гемиплегии, потерей чувствительности на половине тела, потерей зрения на контралатеральной стороне (гемианопсия) и потерей слуха (гемигипоакузия).
Функции таламуса и последствия их нарушении
Таламус играет центральную роль в обработке сенсорной информации поступающей к . Все сенсорные сигналы соматической и других видов чувствительности, за исключением обоняния, проходят к коре через таламус. Как уже упоминалось, сенсорная информация направляется таламусом в кору по трем каналам : в строго специфичные сенсорные области — от специфических ядер, MKT, ЛKT; в ассоциативные области коры — от ассоциативных ядер и ко всей коре — от неспецифических ядер таламуса.
Таламус участвует в частичном восстановлении таких сенсорных ощущений, как болевые, температурные и грубое осязание, которые исчезают после повреждения сенсорной коры. При этом восстановление ощущения боли, сигналы которого передаются волокнами С-типа, проявляется ноющей, жгучей, нс адресованной к какой-либо части тела болью. Предполагают, что центром таких болевых ощущений является таламус, в то время как ощущение острой, хорошо локализованной боли, передаваемой волокнами А-типа, является соматосенсорная кора. Это болевое ощущение исчезает после повреждения или удаления данной области коры.
У больных с острыми нарушениями кровообращения в области таламуса могут развиться признаки таламического синдрома . Одним из его проявлений является потеря всех видов чувствительности на контралатеральной половине тела по отношению к стороне поврежденного таламуса. Однако через некоторое время грубые ощущения боли, осязания и температуры восстанавливаются.
Одной из важнейших функций таламуса является интеграция сенсорной и моторной деятельности . Ее основой является поступление в таламус не только сенсорных, но и сигналов из моторных областей мозжечка, базальных ганглиев, коры. Предполагается, что в вентральном латеральном ядре таламуса локализован треморогенный центр.
Таламус, в котором находится часть нейронов ретикулярной формации ствола мозга, играет центральную роль в поддержании сознания и внимания. При этом его роль в осуществлении реакций активации и пробуждения реализуется при участии холинергических, серотонинергических, норадренергических и гнетаминергических нейромедиаторных систем, которые начинаются в стволе мозга (ядро шва, голубоватое пятно), основании переднего мозга или гипоталамусе.
Через связи медиального таламуса с прсфронтальной корой таламус участвует в формировании аффективного поведения. Удаление префронтальной коры или ее связей с дорзомедальным ядром таламуса вызывает изменения личности, характеризующиеся потерей инициативы, вялостью аффективной реакции, индифферентностью к боли.
Через связи передних таламических и других ядер таламуса с гипоталамусом и лимбическими структурами мозга обеспечивается их участие в механизмах памяти, контроля висцеральных функций, эмоционального поведения. При заболеваниях таламуса могут развиться различные типы нарушений памяти от мягкой забывчивости с рассеянностью до выраженной амнезии.
Оглавление темы "Передний мозг, prosencephalon. Промежуточный мозг, diencephalon.":Таламус, thalamus. Строение таламуса. Ядра таламуса. Функции и значение таламуса
Thalamus, таламус, представляет собой большое парное скопление серого вещества в боковых стенках промежуточного мозга по бокам III желудочка , имеющее яйцевидную форму, причем передний его конец заострен в виде tuberculum anterius , а задний расширен и утолщен в виде подушки, pulvinar .
Деление на передний конец и подушку соответствует функциональному делению thalamus на центры афферентных путей (передний конец) и на зрительный центр (задний) .
Дорсальная поверхность покрыта тонким слоем белого вещества - stratum zonule . В латеральном своем отделе она обращена в полость бокового желудочка, отделяясь от соседнего с ней хвостатого ядра пограничной бороздкой, sulcus terminalis, являющейся границей между telencephalon , к которому принадлежит хвостатое ядро, и diencephalon , к которому относится таламус . По этой бороздке проходит полоска мозгового вещества, stria terminalis .
Медиальная поверхность таламуса , покрытая тонким слоем серого вещества, расположена вертикально и обращена в полость III желудочка, образуя его латеральную стенку. Сверху она отграничивается от дорсальной поверхности посредством белой мозговой полоски, stria medullaris thalami . Обе медиальные поверхности таламусов соединены между собой серой спайкой - adhesio interthalamica , лежащей почти посередине. Латеральная поверхность таламуса граничит с внутренней капсулой, capsula interna .
Нижней своей поверхностью таламус располагается над ножкой мозга, срастаясь с ее покрышкой. Как видно на разрезах, серая масса таламуса белыми прослойками, laminae medullares thalami , разделяется на отдельные ядра, носящие названия в зависимости от их топографии: передние, центральные, медиальные, латеральные, вентральные и задние.
Функциональное значение таламуса очень велико. В нем переключаются афферентные пути: в его подушке, pulvinar , где находится заднее ядро, оканчивается часть волокон зрительного тракта (подкорковый центр зрения, ассоциативное ядро таламуса), в передних ядрах - пучок, идущий от corpora mamillaria и связывающий таламус с обонятельной сферой, и, наконец, все остальные афферентные чувствительные пути от нижележащих отделов центральной нервной системы в остальных его ядрах, причем lemniscus medialis заканчивается в латеральных ядрах.

Таким образом, thalamus является подкорковым центром почти всех видов чувствительности . Отсюда чувствительные пути идут частью в подкорковые ядра (благодаря чему таламус является чувствительным центром экстрапирамидной системы), частью - непосредственно в кору (tractus thalamocorticalis) .




